
Figure 43. Blake Nose, NW Atlantic Sites 1049a and C and Site 1050C. All have similar lithologies, except for the absence of a spherule layer in Site 1050C. Note the highly irregular contact at the top and bottom of the spherule layer an the mottled bioturbated carbonate ooze. From Keller et al., 2013.
"Unequivocal proof that Chicxulub is KTB age"
"First-order agreement with prediction of impact hypothesis Biostratigraphy and Age Control"
Biostratigraphy and Age Control
The strongest age claims for a KTB age for the Chicxulub impact are based on a select few deep-sea localities where a single thin impact spherule layer is reported directly between the Cretaceous and Paleocene. These localities include the Bass River Core of New Jersey, NW Atlantic Site 1049, Caribbean Site 1001 and SW Atlantic Site 1259. (Olsson et al., 1997; Sigurdsson et al., 1997; Norris et al., 1999, 2000; Klaus et al., 2000; Martinez-Ruiz et al., 2001; Huber et al., 2002; MacLeod et al., 2007). This single spherule layer is cited as the ultimate and unequivocal proof that the Chicxulub impact caused the KTB mass extinction. For example, MacLeod et al. (2007) argued that the 2-cm-thick spherule layer that underlies early Danian sediments is in “first-order agreement with the prediction of the impact hypothesis” recording history “within minutes of the impact.” This conclusion was reached despite the evidence of soft- sediment deformation and erosional contacts between the spherule layer, chalk, and white clay at Demerara Site 1259. And no high-resolution biostratigraphic, quantitative faunal and chemostratigraphic analyses that could evaluate the completeness of the KTB transition have been published for any of these sites.
Blake Nose: We analyzed two holes at Site 1049 (A, C) and Site 1050C in order to insure replicable results and exclude artifacts of sampling or processing. All three sites analyzed show very similar lithologies, except for the absence of a spherule layer at Site 1050C (Fig. 43).
Upper Maastrichtian sediments at Sites 1049A and 1049C consist of disturbed, bioturbated, mottled light and darker gray nannofossil and foraminiferal ooze, which underlie the 10-15 cm thick spherule layer (Fig. 43). The spherule layer has a carbonate ooze matrix, irregular bedding, clasts of limestone, chalk and dolomite and is bounded by irregular surfaces. At Site 1049A a 2 mm thin red-brown limonite layer and 8 cm thick white ooze overlie the spherule layer followed by gray carbonate ooze. At Site 1049C, the limonite layer is also present but discontinuous and white ooze is present in the middle of the spherule layer. Above the limonite layer the 8 cm dark and light gray ooze is strongly mottled and bioturbated, followed by light gray ooze. At Site 1050C the KTB transition consists of dark and light gray burrowed ooze with some large horizontal burrows infilled with gray ooze that contains rare Danian species below the KTB (Fig. 43). An erosional surface and darker gray layer is correlative with the spherule layer.
Upper Maastrichtian faunal assemblages at all three sites are diverse, abundant and well preserved but lack Plummerita hantkeninoides, the zone CF1 index species for the topmost Maastrichtian (Figs. 44-46). However, the presence of Gansserina gansseri (and G. wiedenmayeri in Site 1050C) and Pseudoguembelina hariaensis mark this interval as zone CF3 (Abramovich et al., 2010). Micula prinsii was reported from a short interval below the spherule layer (Norris et al., 1999), which is consistent with the upper part of zone CF3 (Fig. 2). This indicates a major hiatus that spans Maastrichtian zones CF1 and CF2 (~ 280 kyr, time scale of Gradstein et al., 2004, Fig. 47). (Keller et al., 2013).
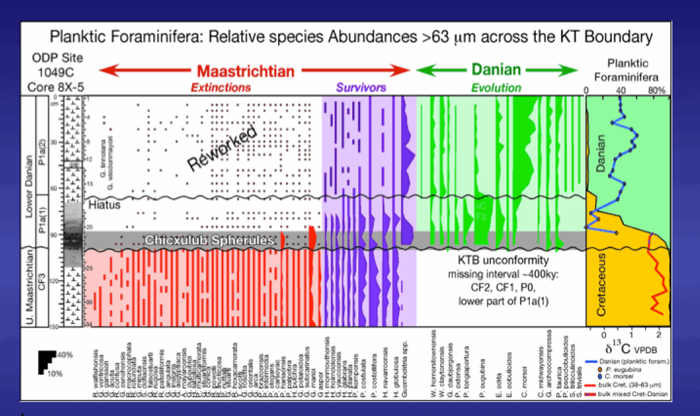
Figure 44. Biostratigraphy, species ranges and species abundances and stable isotopes across the KTB at ODP Site 1049A. Note the abundance and high diversity (12 species) of early Danian species in the spherule layer. Such high species diversity is not seen for at least 100 ky after the mass extinction. From Keller et al., 2013.
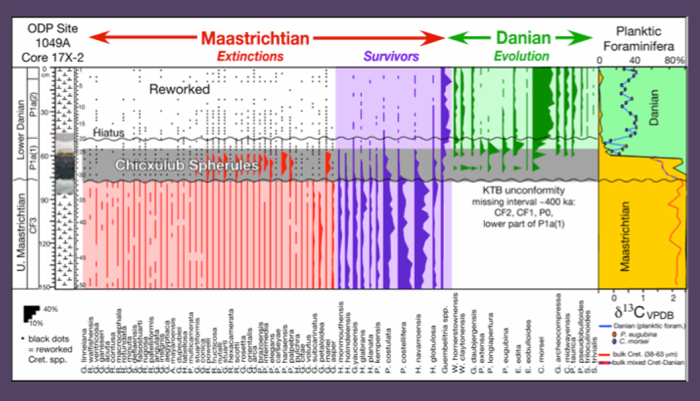
Figure 45. Biostratigraphy, species ranges and species abundances and stable isotopes across the KTB at ODP Site 1049A. Note the abundance of reworked Cretaceous species in the spherule layer, but also significant abundance of early Danian species that indicates spherule redeposition in the early Danian. A KTB unconformity marks a missing interval of about 400 ky with zones CF2, CF1, P0 and the lower part of P1a(1) missing. Cretaceous reworked species decrease in abundance above the spherule layer. From Keller et al. 2013.
The spherule-rich layer above the KTB unconformity at Sites 1049A and 1049C is 15 cm and 10 cm thick, respectively (Figs. 44, 45). Reworked Cretaceous species are common along with shallow and deep-water benthic foraminifera that indicate reworking and transport of shallow water sediments into the basin (Alegret and Thomas, 2004). We also observed a dominance of large and thick-shelled species with variable preservation, including discoloration, abrasion and broken shells, and low abundance of small species, all of which indicate reworking and transport. On a quantitative basis 83-95% (>63 mm) of the Maastrichtian species are reworked in the spherule layer and 5-17% are small Danian species. Early Danian species are abundant and diverse with 12 species in the spherule layer. Such high abundance or high diversity is not seen in the early Danian until about 100ky after the KTB mass extinction. This indicates that the spherule layer was deposited long after the KTB at a time of reworking and transport of sediments on Blake Nose.
The relative abundance of Danian species was calculated based on the 63-100 mm size fraction from the spherule layer. The presence of diverse assemblages (12 species), including P. eugubina and P. longiapertura, but absence of P. pseudobulloides and/or S. triloculinoides indicates a subzone P1a(1) assemblage in the spherule layer with the lower part of this subzone and zone P0 missing. A similar Danian assemblage is present in the 8 cm above the spherule layer at Site 1049A and in the 20 cm above the spherule layer at Site 1049C, whereas the abundance of reworked Cretaceous species rapidly decreases to <10% (Fig. 45). The first appearances P. pseudobulloides, S. triloculinoides and the sharp increase in Chiloguembelina morsei and Guembelitria species 8 cm and 20 cm above the spherule layer at Sites 1049A and 1049C marks a hiatus between P1a(1) and P1a(2). At both sites P1a(2) is dominated by two opportunistic species, C. morsei and G. cretacea, that indicate increased environmental stress. Reworked Cretaceous species are still present but rare (<2%). Keller et al., 2013.

Figure 46. Illustration of the KTB criteria and age assigned by Norris et al., 1999 at Site 1049A. Large red circles mark the KTB localities analyzed by Keller et al., 2013 to test the assumption that the Chicxulub is precisely KTB in age.
Earlier studies of Site 1049A placed the KTB at the base of the spherule layer based on the assumption that the Chicxulub impact is KTB age (Norris et al., 1999; Schulte et al., 2010), and supporting this assumption by “a 10 cm thick spherule layer precisely at the KTB … demonstrates that the impact was synchronous with the mass extinction.” The age of the underlying Maastrichtian carbonate ooze was assumed to be the latest Maastrichtian age with no unconformity present (Fig. 46). This age interpretation was based on the presence of a short interval of M. prinsii zone, which spans more than planktic foraminiferal zones CF1 and CF2 into CF3, and Abathomphalus mayaroensis, which spans the late Maastrichtian zones CF4 to CF1. The absence of zone CF1 (absence of P. hantkeninoides) was explained as absent in this part of the N. Atlantic. But in fact this species is present in the N. Atlantic, Caribbean (Site 1259, Demerara), Cuba, Gulf of Mexico, NE Mexico and Texas. The absence of zone CF1 at Site 1049 should have been the warning signal that this section is incomplete. Compounding this error is the placement of the first appearance of P. eugubina at the top of the spherule layer. No mention was ever made of the common occurrence of Danian species through the spherule layer. These errors led to a string of wrong conclusion, namely that the KTB on Blake Nose is complete, that the impact spherules are precisely synchronous with at the KTB mass extinction and that therefore proof that the Chicxulub impact is precisely KTB age.
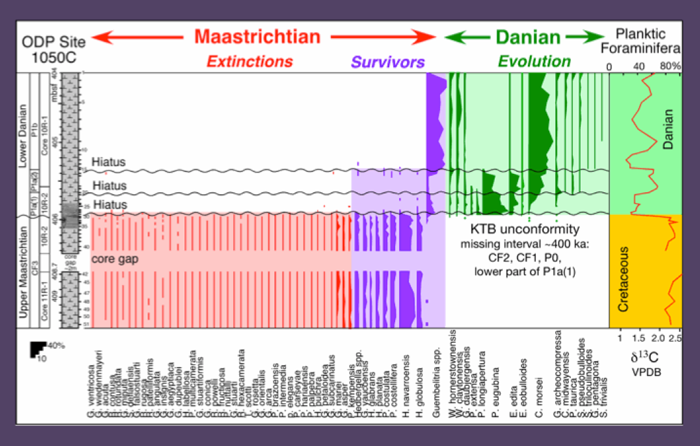
Figure 47. Biostratigraphy, species ranges and species abundances and stable isotopes across the KTB at ODP Site 1050C. The hiatus at this locality is of the same magnitude as at Site 1049, but no impact spherules are present. This is likely because Site 1050 is located at a distance from the submarine channels that supplied the impact debris to Site 1049. This is also indicated by the absence of significant reworking of Cretaceous species at or above the KTB. From Keller et al., 2013.
At Site 1050C the comparable P1a(1) and P1a(2) assemblages spans the 60 cm above the KTB unconformity. Reworked species are very rare (Fig. 47). The basal Danian sample is dominated by P. eugubina (50%), which indicates that zone P0 and the lower part of P1a(1) is missing along with zones CF1 and CF2 (~400 kyr). Subzone P1a(1) is dominated by P. eugubina (55-64%) with common Eoglobigerina edita and variable presence of seven other Danian species. Short hiatuses are also present at the P1a(1)/P1a(2) and P1a(2)/P1b boundaries (see Keller et al., 2013).
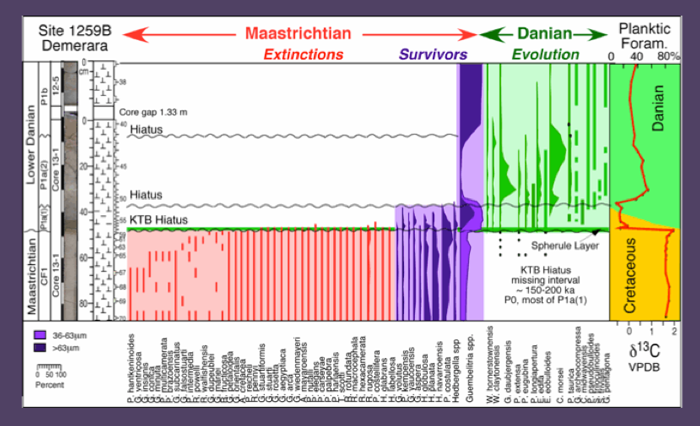
Figure 48. Biostratigraphy, species ranges and species abundances and stable isotopes across the KTB at Demerara ODP Site 1250B. The spherule layer is only 2 cm thick at this location. The latest Maastrichtian zone CF1 was identified though a short KTB hiatus is present along with short hiatuses at the P1a(1)/P1a(2) and P1a(2)/P1b zone boundaries. From Keller et al., 2013.
Demerara Rise: Sediments below the 2 cm thick spherule layer consist of mottled, bioturbated marly chalk with a relatively sharp upper contact. The spherule layer is graded and thin white chalk layers are present at the base and top, similar to Site 1001B. Danian sediments consist of strongly mottled, bioturbated light and darker gray clayey chalk.
Preservation of planktic foraminifera is good for the Danian and moderate in the Maastrichtian with dissolution effects (thin walls, holes in test chambers). The marly chalk below the spherule layer contains a diverse late Maastrichtian planktic foraminiferal assemblage, including rare Plummerita hantkeninoides, the index species for zone CF1 (Fig. 48). Cretaceous assemblages in the >63 µm are dominated by small biserial species (Pseudoguembelina costulata, Heterohelix planata, H. navarroensis), whereas Guembelitria cretacea is common in the smaller size fraction (36-63 mm). Rare Danian species were obtained from the 10 cm thick bioturbated light and dark grey clay below the KTB unconformity and spherule layer; these specimens are likely due to burrowing from the overlying Danian. Keller et al., 2013.
One sample was obtained from the spherule layer. Upper Maastrichtian planktic foraminifera are common in the >63 mm and small Danian species are common in the 36-63 mm size fraction where Guembelitria species dominate (83-96%) in the spherule layer and up to 8 cm above it (Fig. 48). The presence of P. eugubina, G. daubjergensis, W. hornerstownensis, W. claytonensis and E. eobulloides in the spherule layer and the 8 cm above indicate subzone P1a(1) (Keller et al., 2013).
Stable isotopes: For Blake Nose (Sites 1049A, 1049B and 1050) and Demerara Site 1259B stable isotopes we analyzed bulk rock fine fraction (36-63mm), which consists primarily of nannoplankton and small planktic foraminifera and is generally a good indicator of primary productivity (Zachos et al., 1989; Schrag et al., 1995). For the Maastrichtian d13C values vary between 1.8-2.2‰, except for Site 1050C which is slightly heavier (2.2-2.4‰, Fig. 48). These values are comparable to those observed in open marine environments, such as South Atlantic Site 525A (Li and Keller, 1998a). For Blake Nose and Demerara no significant changes in d13C are apparent for the intervals of CF3 and CF1 analyzed below the KTB unconformity. For Blake Nose this is due to the unconformity with the interval of zones CF2 and CF1 missing; where the major climatic changes occurred. Keller et al., 2013.
Age and Origin of Impact Spherules
A KTB age for the Chicxulub impact was assumed based on spherules overlying Maastrichtian sediments at the five sites investigated (e.g., Olsson et al., 1997; Sigurdsson et al., 1997; Norris et al., 1999; MacLeod et al., 2007). There are three key factors that argue against this interpretation:
i) major KTB unconformity in all sequences, though erosion is less extensive at Site 1259B;
ii) spherule-rich sediments contain common reworked Maastrichtian species and shallow water benthic foraminifera (Alegret and Thomas, 2004), which argues for erosion and redeposition,
ii) spherule-rich layer contains a diverse early Danian P1a(1) species assemblage, which indicates deposition during the early Danian well after the mass extinction and evolution of the first Danian species.
Earlier studies reported the first Danian species above the spherule layer and thereby justified placing the KTB at its base. It is possible that the early Danian assemblages were not observed because of their small size and investigations were limited to the >63 microns size fraction.
One might argue that the Danian species in the spherule layer are an artifact of bioturbation or leaking of sediments into the more porous spherule layer. Some bioturbation is clearly evident in the lithology of Sites 1049A, 1049C, 1050C (Fig. 43). Similarly at Site 1259B, rare Danian species were observed in burrows below the KTB unconformity, contrary to Schulte et al. (2009, p. 1185) who noted “burrows or other signs of bioturbation are totally lacking “. Although bioturbation may account for some Danian specimens in the spherule layer, this could not account for the diverse assemblage with hundreds of specimens and similar species abundances and diversity as in assemblages above the spherule layer.
The presence of these Danian assemblages in the spherule layer above the unconformity indicate that sedimentation resumed in the upper subzone P1a(1) about 150-200 kyr after the KTB mass extinction. This indicates that the spherules are eroded, transported and redeposited long after the Chicxulub impact. Because the normal early Danian evolutionary pattern is present, except for the missing basal Danian, we can assume that the Danian fauna represents in situ deposition but with added downslope transport of spherule-rich reworked debris and Maastrichtian foraminifera. The absence of spherules in Site 1050C can be explained by its distant location from the submarine channel that transported debris to Sites 1049. Similar reworking of shallow water sediments and spherules, transport and redeposition into deeper waters is also observed in NE Mexico sections and Site 1259 (Alegret et al., 2001, 2002; Alegret and Thomas, 2004). Based on the results from this study the spherules in lower Danian sediments must be reworked from an older primary deposit. Reworked impact spherules have previously been observed in Danian sediments from Haiti, Cuba, Belize, Guatemala and southern Mexico (Keller et al., 2001, 2003b; Alegret et al., 2005; Arenillas et al., 2006; Keller, 2008).
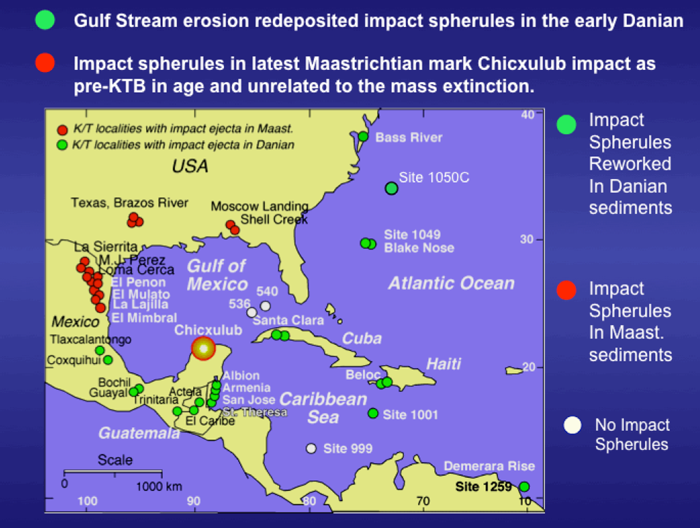
Figure 49. Location of KTB sites analyzed by our team in the area surrounding the Chicxulub impact. Note that KTB localities with impact spherules in the latest Maastrichtian are in NE Mexico and Texas. KTB localities with impact spherules reworked into early Danian P1a sediments are in locations from southern Mexico, Guatemala, Belize through the Caribbean and North Atlantic including Cuba and Haiti. (From Keller et al., 2013).
KTB Unconformity
A thin impact spherule layer between Maastrichtian and Paleocene sediments in some KTB sequences from the North Atlantic and Caribbean is commonly cited as proof that the Chicxulub impact is KTB in age and the sedimentation record complete (e.g., Olsson et al., 1997; Sigurdsson et al., 1997; Norris et al., 1998, 1999; Martinez-Ruiz et al., 2000; MacLeod et al., 2007; Schulte et al., 2010).
Results based on high-resolution biostratigraphy, quantitative faunal analyses and stable isotopes show that far from being complete all seven sections analyzed show a major unconformity across the KTB with variable erosion of Maastrichtian sediments. In all sections where impact spherules are present they are reworked into early Danian sediments tens of thousands of years after the KTB. Clearly, these impact spherules in Danian sediments above the KTB unconformity cannot be taken as proof that the Chicxulub impact hit Yucatan precisely at KTB time.
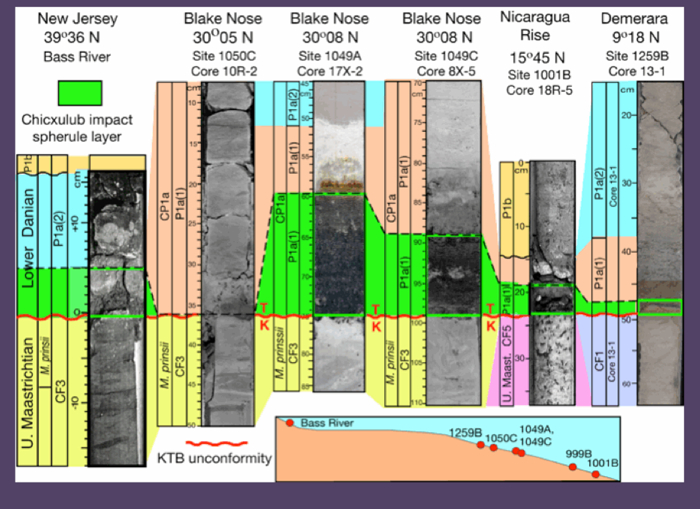
Figure 50. KTB unconformity from the North Atlantic through the Caribbean. At all locations where Chicxulub impact spherules are present they are reworked into early Danian sediments tens of thousands of years after the KTB mass extinction. (Keller et al., 2013).
The maximum extent of erosion can be estimated based on Paleocene and Maastrichtian sediments above and below the unconformity, respectively. At Bass River, New Jersey, the KTB unconformity spans from the early Danian subzone P1a(2) through the upper Maastrichtian zones CF1 and CF2 (~620 ky), in North Atlantic Sites 1049A, 1049C and 1050C from P1a(1) through CF1 and CF2 (~400 kyr, Figs. 45-48), and in the Caribbean Sites 999B and 1001B from P1a(1) through zones CF1, CF2, CF3 and CF4 (~3 myr). Only at Demerara Rise Site 1259B is erosion or non-deposition more limited spanning most of subzone P1a(1) and zone P0 (~150-200 kyr, Fig. 48). The nearby Site 1258A provides some clues to possible erosion at the top of the Maastrichtian. Just below the KTB Thibault and Gardin (2006) record the abrupt end of the nannofossils Micula murus acme that marks the CF1 warm event. In complete sequences, this global warm event ends with the onset of cooling about 50-100 kyr prior to the KTB (Li and Keller, 1998a; Abramovich and Keller, 2003; Abramovich et al., 2011), which suggests erosion of the latest Maastrichtian at Site 1258A and likely at Site 1259B (Keller et al., 2013).
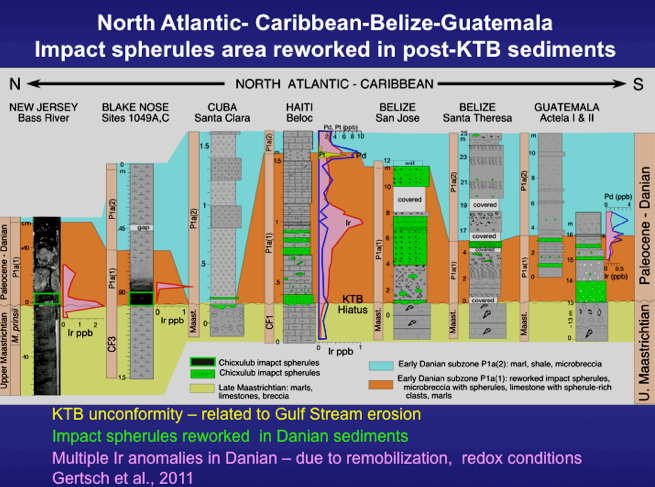
Figure 51. Impact spherule distribution from the North Atlantic through the Caribbean, including Cuba, Haiti, Belize and Guatemala reveal impact spherules reworked into early Danian zone P1a sediments with an Ir anomaly at the top of the spherule layers.
One might argue that the KTB unconformity New Jersey, Blake Nose to Demerara Rise is unusual and must have an extraordinary cause, such as the Chicxulub impact (e.g., Bralower et al., 1998; Arenillas et al., 2006; Schulte et al., 2010). However, this unconformity is not unique and is documented from all localities in the NW-SW Atlantic, Caribbean and Gulf of Mexico, Cuba, Haiti, Belize, Guatemala and southern Mexico (Keller et al., 2003). Moreover, major unconformities are the norm in this region through the Cretaceous (Keller et al., 1993). Most workers concentrated on the KTB and may have been unaware of this active erosion likely due to the early Gulf Stream current, or believed strongly that only the Chicxulub impact could have caused it. In addition, small Ir anomalies are frequently found above the reworked Danian spherule layers, similar to Bass River, Blake Nose and Demerara. Keller et al. (2003) suggested that these Ir anomalies indicate a second impact after the KTB. However, since the early 2000 much has been learned about Ir concentrations, redox boundaries and re-mobilization of Ir well after deposition (e.g., Miller et al., 2010, Martin- Peinado and Rodriguez-Tovar, 2010; Gertsch et al., 2011; Premovic and Ilic, 2012). The source of these Ir anomalies is likely this secondary process and no impact origin has to be invoked.
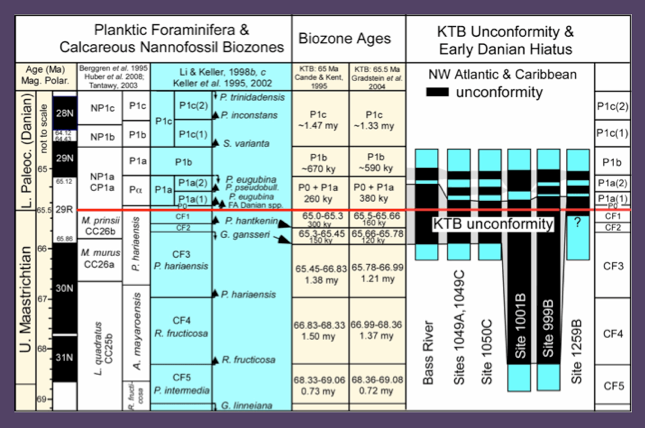
Figure 52. Biostratigraphy and ages for biozones based on the KTB at 65 Ma and 65.5 Ma. The extent of the KTB unconformity in the N. Atlantic and Caribbean as well as the shorter early Dainian hiatues are indicated. From Keller et al., 2013.
Shorter pulses of erosion in the early Danian, as documented in this study, are also widespread (MacLeod and Keller, 1991a,b; Keller et al., 2003a, this study). Moreover, impact spherules and frequently small Ir anomalies are present above the KTB unconformity in early Danian P1a sediments in Cuba and southern Mexico (Alegret et al., 2005; Arenillas et al., 2006), in Haiti, Belize, and Guatemala (Keller et al., 2001, 2003b; Keller, 2008) and in the Caribbean and N. Atlantic (this study). Impact spherules are abundant and restricted to the latest Maastrichtian zone CF1 (~130-150 kyr below KTB) in NE Mexico and Texas where KTB sequences are more complete. Their absence below the KTB in the Atlantic, Caribbean, Gulf of Mexico, southern Mexico, Haiti, Belize and Guatemala is likely due to erosion linked to intensified current circulation and the impact (Keller et al., 1993; Watkins and Self-Trail, 2005).
Bralower et al. (1998) attributed any reworked microfossils, impact derived materials, lithic fragments and unconformities in the Caribbean and Gulf of Mexico to the Chicxulub impact and proposed that such deposits (labeled impact cocktail) are a reliable way to identify the Cretaceous-Tertiary boundary. All of the sections discussed in their paper show faunal and hiatus patterns similar to the Atlantic and Caribbean sections of this study where impact spherules are reworked in the early Danian and provide no evidence for a KTB age for the Chicxulub impact.
Thus, caution must be exercised in interpreting Ir anomalies and impact spherules as KTB age markers for the Chicxulub impact, limestone conglomerates and breccias as related to the Chicxulub impact, and unconformities and hiatuses as caused by this impact. Sediment depositional patterns, impact spherule layers, minor Ir anomalies, short hiatuses and major unconformities through N. Atlantic, Caribbean, Gulf of Mexico and Central America are far too variable and complex for a simple one-size-fits-all interpretation.