
Figure 21. Location of Jhilmili and the Jhilmili secton. From Keller et al., 2009a.
Deccan intertrappean sediments in central India are generally terrestrial deposits of Maastrichtian age. However, we discovered the KTB transition in a marine incursion within intertrappean sediments at Jhilmili, Chhindwara District, Madhya Pradesh. Integrative biostratigraphic, sedimentologic, mineralogic and chemostratigraphic analyses reveal the basal Danian in the intertrappean sediments between lower and upper trap basalts that regionally correspond to C29r and the C29R/C29N transition, respectively. From Keller et al., 2009a.

Figure 22. Jhilmili, Chhindwara, central India, intertrappean sediments between Phase-2 and Phase-3 sediments reveal deposition in a lacustrine-brackish environment in the earliest Danian. From Keller et al., 2009a.
Lithology and Paleoenvironments
The Jhilmili intertrappean sediments span nearly 14 m between lower and upper basalt flows (Fig. 22). Lithologies range from claystones (and paleosols), siltstones, clayey–silty marlstones to limestones and can be subdivided into 5 different units, including the lower and upper traps (Fig. 23). These units are described in Keller et al., 2009a and illustrated in Fig. 23. Of importance here is unit 3, the interval where planktic foraminifera and rare brackish water ostracods were found. Their presence indicates deposition in fresh water to brackish-marine conditions probably due to a marine incursion (Keller et al., 2009a).

Figure 23. Lithology, micrographs and paleoenvironmental characteristics of the Jhilmili sequence. Micrographs: 1-2, Paleosol of upper unit 5 with root molds (d), charcoal fragments (a), glass spherules (b) and shards (c). 3-4, Stromatolitic microbial mat with embedded ostracod (3) and a charophyte gyrogonite (4). 5-6, Stromatolitic microbial mats showing typical tepee features, indicating emersion. 7-10, Volcanic glass rich layer (sample JH18). From Keller et al., 2009a.

Figure 24. Clay mineral composition, XRD diffractograms and paleoclimatic interpretation. Note the marine incursion marks the transition from subhumid to more arid climate conditions, though wet and dry seasonal cycles continue. (Keller et al., 2009a)
Clay mineral assemblages reflect continental morphology, tectonic activity, climate changes and sea level fluctuations and are therefore excellent environmental proxies. At Jhilmili, clay mineral patterns reveal distinct climatic and environmental changes (Fig. 24). At the base the 100% smectite indicates a typical paleosol derived mainly from weathering of basaltic rocks in semi-arid seasonal climate with alternating wet and dry seasons. The appearance of palygorskite in unit 3 indicates increasingly dry conditions in the source area. Palygorskite may form along coastal and peri-marine environments where continental alkaline waters are concentrated by evaporation forming Si-rich brines, which favor precipitation of palygorskite and/or smectite (Pletsch, 1996). Similar climate conditions have been inferred for upper Maastrichtian intertrappean sediments from Anjar in which palygorskite and smectite are dominant (Khadkikar et al., 1999, Shrivastava and Mansoor, 2005) and intertrappean beds at Rajahmundry (Keller et al., 2008). See Keller et al., 2009a). The poorly crystallized illite in unit 4 may have derived from a partial degradation of smectite from lacustrine environments (Deconninck et al., 1988; Singer and Stoffers, 1980). In unit 5 the abundant and poorly crystallized illite (up to 80%, Fig. 24) is likely to have derived from the transformation of smectite due to increased temperature resulting from deposition of the upper lava flow (unit 6).
Biostratigraphy
The discovery of planktic foraminifera in the Jhilmili section was fortuitous. Routine analysis of ostracods by two independent Indian research groups detected foraminifera. Follow-up collection and analysis by Keller and Adatte revealed a terrestrial sequence between phase-2 and phase-3 lava flows with a thin interval of lacustrine to brackish marine sediments with planktic foraminifera equivalent to the early Danian zone P1a (Fig. 16, 25) (Keller et al., 2009a, b).
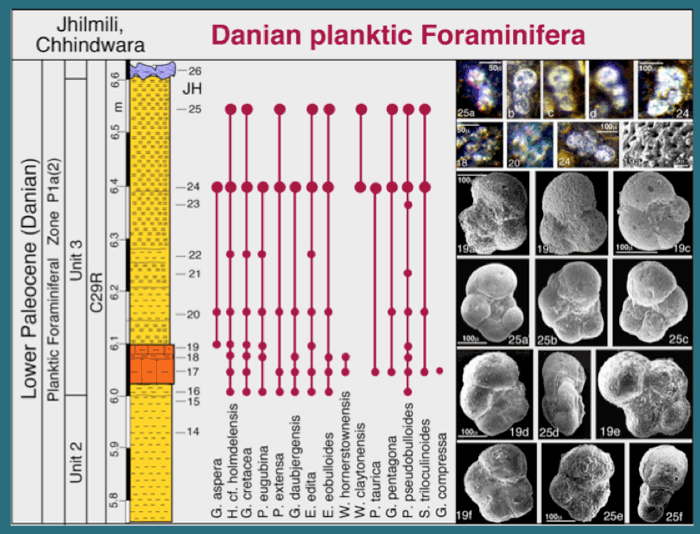
Figure 25. Jhilmili, Planktic foraminiferal biostratigraphy and species ranges of Jhilmili intertrappean sediments. Larger circles mark samples with common foraminifera. Small planktic foraminiferal species are illustrated from thin section micrographs. Scale bar is the same for each row, unless otherwise indicated; numbers refer to samples. 24, 25a-d, Parasubbotina pseudobulloides; 18, 19, Parvularugoglobigerina eugubina; 19a, SEM of cancellate spinose wall texture with calcite overgrowth of chambers in Subbotina triloculinoides 19a. 19b, 19c, Subbotina triloculinoides; 25a-c, 19d, P. pseudobulloides; 19e, 25d, Hedbergella cf. holmdelensis; 19f, G. pentagona; 25e, Subbotina triloculinoides; 25f, Globigerinelloides aspera. From Keller et al., 2009a,b.
Biostratigraphic analysis is based on planktic foraminifera, which permit high-resolution age control and correlation to the marine record. Nearly all samples in unit 3 contain planktic foraminifera in thin sections and/or washed residues. Thin section assemblages consist predominantly of small (40-100 µm), poorly preserved, earliest Danian species, including Parvularugoglobigerina eugubina, P. extensa, Globoconusa daubjergensis, Eoglobigerina edita, E. eobulloides and Woodringina hornerstownensis. Larger specimens (>150µm) from washed residues include early Danian species Parasubbotina pseudobulloides, Subbotina triloculinoides, Praemurica taurica and Globigerina (E.) pentagona (Fig. 25). Also present are Cretaceous species Globigerinelloides aspera, Hedbergella cf. holmdelensis and disaster opportunist Guembelitria cretacea, which are generally considered KTB survivors (Keller et al. 2002; Keller et al., 2009). This assemblage is typical of the early Danian P. eugubina zone P1a globally and a similar assemblage has been identified from intertrappean beds of the Rajahamundry quarries (Keller et al., 2008).
In these Jhilmili intertrappean sediments the presence of zone P1a index species indicates that the foraminifera-bearing interval was deposited about 100-150 ky after the K-T boundary in subzone P1a(2) (Fig. 17). The underlying 6 m of red shale in unit 2 consist mainly of paleosols and were likely deposited during the earlier Danian P0-P1a(1) interval, as suggested by the presence of this interval in intertrappeans of Rajahmundry (Keller et al. 2008). The upper red and green shales of units 4 and 5 represent paleosols and palustrine conditions and were likely deposited during subzone P1a(2), which precedes the C29r/C29n transition. The Jhilmili intertrappean sediments thus record the age and environmental conditions following the K-T mass extinction, which appears to have coincided with the end of the main phase of Deccan volcanism (Ambenali Formation). From Keller et al., 2009a.
Paleoenvironment – Microfossils and Microfacies
Microfossils and microfacies provide clues to the depositional environment at Jhilmili. Ostracods and algae (charophytes) are common to abundant (though not age diagnostic), with planktic foraminifera variable in unit 3. Nearly all ostracod species are known from fresh water deposits (Whatley and Bajpai, 2005). Only two species are indicative of brackish water environments: a rare occurrence of Buntonia sp. (sample JH21) and common Neocyprideis raoi (JH21 to JH26) along with common planktic foraminifera. Abundant N. raoi were recorded from intertrappean beds of the east coast of India where they mark brackish deposits (Khosla and Nagori, 2002). This suggests that the brackish ostracods and planktic foraminifera in the middle unit 3 are due to short-term intermittent influx of marine waters (e.g., storms, high tides) and/or erosion of previously deposited brackish marine sediments. The absence of benthic species indicates that stable marine conditions to support bottom dwellers never developed.
Microfacies of the Jhilmili intertrappean sediments (Fig. 26) support this interpretation. Deposition occurred in predominantly terrestrial (paleosol) environments, except for the middle unit 3, which marks a transition to palustrine and fresh water conditions followed by brackish-marine environment. The presence of planktic foraminifera in clay clasts and clays of fresh water deposits, as indicated by ostracods and charophytes, reveals current transport of shallow marine-brackish sediments and paleosol clasts from a nearby source. Unit 3 thus represents a predominantly fresh water environment of lakes, ponds and floodplains with cyclic (seasonal) marine incursions transporting planktic foraminifera and repeated erosion and redeposition. From Keller et al., 2009a.

Figure 26. Microfacies and their depositional settings within the terrestrial to freshwater and marine-brackish environments of the Jhilmili intertrappean sequence. Microfacies MF1 to MF7 characterize terrestrial, palustrine, freshwater and marine-brackish conditions. Micrographs with JH designations mark sample locations. From Keller et al., 2009a.
How did forams get into Central India?
The presence of planktic foraminifera in central India can only be explained by a source of marine waters. Jhilmili is located in central India about 800 km from the present coast to the west and equally distant from the eastern coast (Fig. 21). A marine incursion would therefore have originated from an existing major seaway deep into central India. Jhilmili is located near the easternmost extension of the Tapti rift valley, which forms an arm of the Narmada rift valley. The obvious seaway would therefore have been from the west along the Narmada and Tapti rift valleys (Fig. 27). Alternatively, a seaway could have extended north along the Godavari rift valley, but insufficient data is presently available to explore this option. Both seaways could have operated at the same time (Bande et al., 1981), Sahni (1983) as suggested by the distribution of abundant dinosaur eggs and nests surrounding these two rift valleys (Fig. 27). (Keller et al., 2009a, b, c).
Marine incursions along the Godavari and Narmada rift zones are known from the lower Cretaceous and late Cenomanian to early Turonian Bagh Formation based on marine invertebrates, including ostracods, planktic foraminifera and algae (Chiplonkar and Badve, 1968; Badve and Ghare, 1977). Planktic foraminiferal assemblages from the Chirakhan marl of the Bagh Formation indicate a late Cenomanian to early Turonian age, a time of maximum sea level transgression (Sharma, 1976; Rajshekhar, 1996). However, evidence for a seaway during the late Cretaceous is highly controversial and based largely on sedimentological data (e.g., Singh (1981), but see Tandon et al. (1995), though lagoonal deosits have been identified in the Maastrichtian Lameta beds of the Jabalpur area (Shukla and Srivastava, 2008). From Keller et al., 2009a.
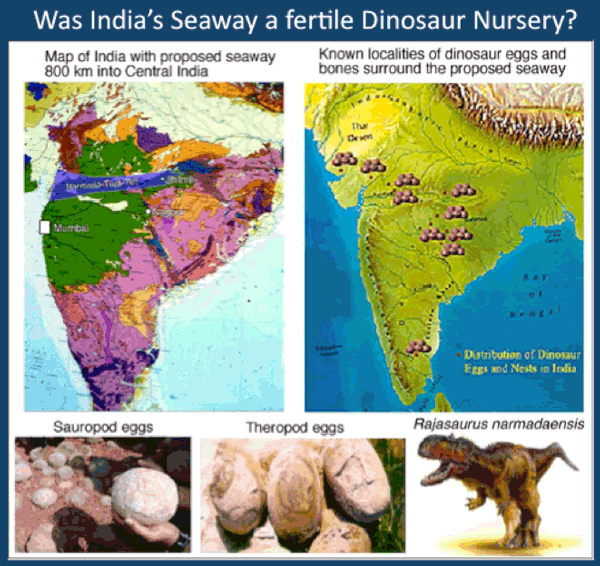
Figure 27. During the Cenomanian a seaway existed from the west into central India through the Narmada and Tapti rift valleys. This seaway is likely to have existed during the KT transition and can explain the presence of marine foraminifera at Jhilmili. The dinosaur egg distribution mirrors the location of this ancient seaway suggesting that this was the location of dinosaur nurseries.
Summary
Intertrappean deposition occurred in predominantly terrestrial semi-humid to arid environments. But a short aquatic interval of fresh water ponds and lakes followed by a shallow brackish marine coastal environment is indicated by ostracods and early Danian zone P1a planktic foraminifera that mark this interval very close to the K-T boundary. This marine incursion marks the existence of a nearby seaway, probably extending inland from the west through the Narmada and Tapti rift valleys. The Jhilmili results thus identify the KT boundary near the end of the main phase of Deccan eruptions, similar to Rajahmundry quarries, and indicate that a major seaway extended at least 800 km across India. This projected seaway is surrounded by abundant dinosaur nesting sites and appears to have supported thriving dinosaur nurseries. Dinosaurs disappeared with the onset of phase-2 Deccan volcanism.

Figure 28. Death of dinosaurs as a result of Deccan volcanism. Ilustration by NSF artist based on research on Deccan Volcanism by Keller’s team. Fissure eruptions, similar to the Laki eruptions, are shown as the likely type of volcanism.